植物の精細胞を覆う膜の統一名称を決定



横浜市立大学 木原生物学研究所 丸山大輔准教授と、フランス国立農業・食料環境研究所(INRAE、リヨン高等師範学校所属)Thomas Widiez科学ディレクター(Scientific Director)は、世界の主だった専門家たちとともに議論を重ね、このたび、精細胞を覆う膜を「peri-germ cell membrane」と命名しました。本名称は日本語で「生殖細胞を覆う膜」を意味します。
学問において、専門用語は円滑な教育研究活動の基盤となる必須の道具です。以降、世界中の教科書や論文で本名称が使われることからも、今回の名称統一は植物科学の重要な過程といえます。今後、日本語の新名称が専門家の協議で定められる予定です。
本成果は、植物科学誌「Nature Plants」に掲載されました(2024年10月15日)。
研究成果のポイント
精細胞を覆う生体膜に複数の呼称があることで植物生殖研究に混乱が生じていた。
世界中の研究者との協議で精細胞を覆う膜をperi-germ cell membraneと命名。
名称を統一したことで今後の重複受精の研究と教育に重要な貢献をした。
[画像1]https://digitalpr.jp/simg/1706/97072/450_248_20241016141217670f4b3170c37.jpg
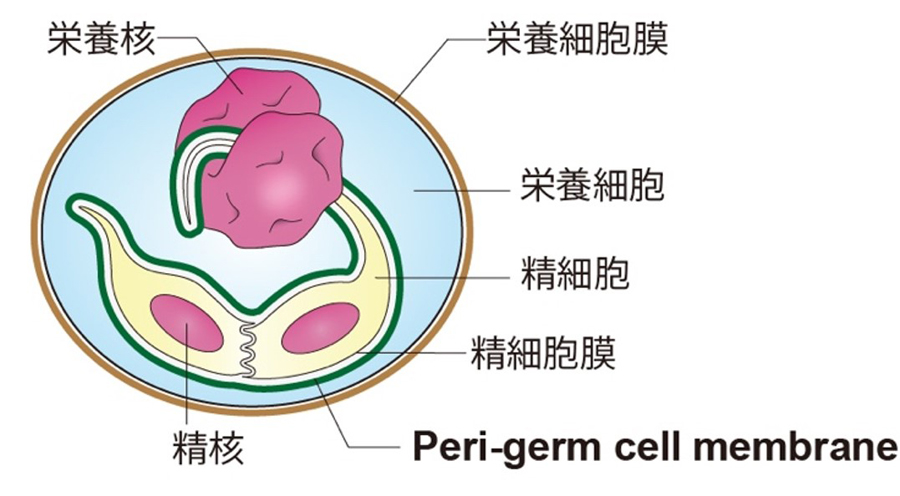
図1 三細胞性花粉の模式図
これまで、文献によって精細胞の外側を覆う膜の呼びかたが異なっていたが、今回、世界の専門家の同意を得て「Peri-germ cell membrane」という名称に統一した
研究背景
植物の種子形成は、2つの精細胞が卵細胞と中央細胞という雌の細胞を同時に受精させる重複受精によって始まります。2つの精細胞は花粉や花粉管の栄養細胞の内部で、一重膜に覆われた状態で存在します(図1)。この膜は重複受精で重要な役割を担っているにも関わらず、統一された名称をもたず、文献によってさまざまな形で呼ばれてきました。
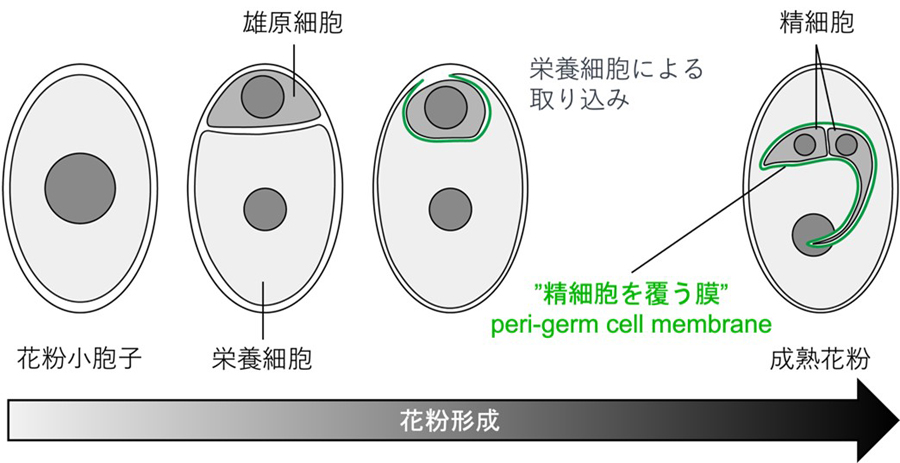
シロイヌナズナやイネ、トウモロコシなどの植物(被子植物)で見られる三細胞性の花粉*1は、雄しべの葯(やく)で小胞子が2回の分裂をすることで作られます。まず、最初の細胞分裂で小さめの雄原細胞と栄養細胞が作られると、栄養細胞は雄原細胞を中に取り込みます。続く細胞分裂で雄原細胞から2個の精細胞が作られて花粉は成熟します(図2)。細胞の取り込みという特徴的な過程を経るため、雄原細胞や精細胞は栄養細胞の細胞膜に由来する一重膜に包み込まれた状態で存在します。この膜は古くから知られており、1969年にはすでに雄原細胞を覆う膜を意味するgenerative cell envelopeと記述されています[1]。ところがその後、internal plasma membrane of the vegetative cell、inner plasma membrane of the pollen grain、inner vegetative cell plasma membrane、pollen endo-plasma membraneなど、この膜はさまざまな名称で呼ばれることとなってきました[2-6]。近年、精細胞を覆う膜が重複受精に果たす役割が議論されるようになり、乱立した専門用語を統一する機運が徐々に高まってきました。
[画像2]https://digitalpr.jp/simg/1706/97072/450_232_20241016141217670f4b31865b6.jpg
図2 三細胞性花粉の花粉形成過程
研究内容
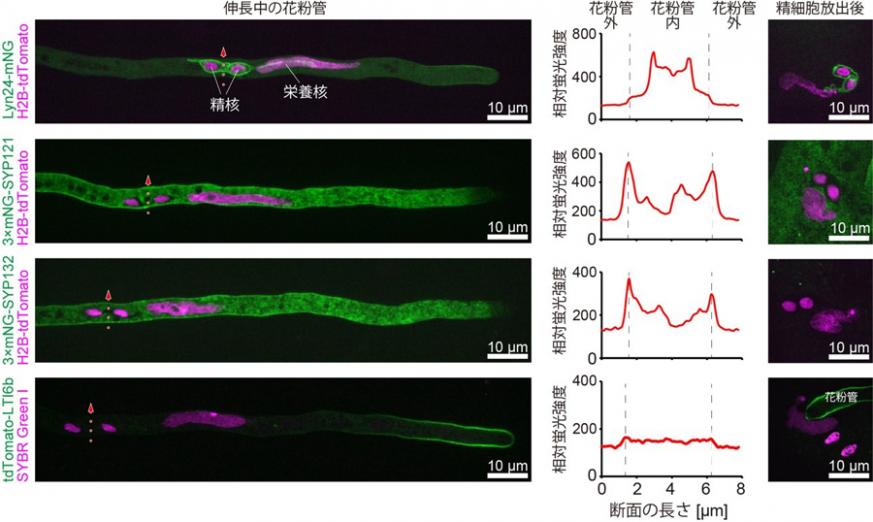
名称の統一に向けては、精細胞を覆う膜の性質を適切に表し、かつ、簡潔で使いやすいものが求められました。しかし、花粉や重複受精をさまざまな観点から扱う世界中の専門家の意見を取り入れ、皆が納得する名称を提案する作業は大きな挑戦でした。そこでThomas Widiez博士と丸山大輔准教授が着目したのは、旧名称のほとんどが細胞膜(原形質膜)を意味するplasma membraneを含む点でした。精細胞を覆う膜は確かに栄養細胞の細胞膜を起源としていますが、近年の研究でこの膜に含まれるリン脂質やタンパク質の構成が通常の細胞膜とは異なることがわかってきています[7-10]。そこで、典型的な3種の細胞膜タンパク質SYP121、 SYP132、 LTI6bを蛍光タンパク質でラベルして、その局在を確認しました。すると、いずれも花粉管栄養細胞の細胞膜には存在する一方、精細胞を覆う膜では検出されないことが示されました(図3)。これは、今回議論となっている精細胞を覆う膜が典型的な細胞膜とは異なる性質を持つことを意味しています。これにより、新名称から“plasma membrane”を除くことで合意が得られました。その後の議論で、雄原細胞と精細胞の両方の意が含まれるgerm cellという語に対して、「周辺の、覆う」の意をもつ接頭辞のperiを合わせたperi-germ cell membraneが新名称として選ばれました。
[画像3]https://digitalpr.jp/simg/1706/97072/450_269_20241016141230670f4b3e9dada.jpg
図3 典型的な細胞膜タンパク質はperi-germ cell membraneに局在しない
蛍光タンパク質でラベルした典型的な細胞膜タンパク質(SYP121, SYP132, LTI6b)とperi-germ cell membrane局在シグナル(Lyn24)をつないだ蛍光タンパク質を花粉管栄養細胞で発現させた。伸長中の花粉管において典型的な細胞膜タンパク質は花粉管栄養細胞膜への局在を示した。一方で、伸長中の花粉管および精細胞放出後の両方においてperi-germ cell membraneには局在しなかった。赤い矢印で示した部分の相対蛍光強度をグラフに示している
今後の展開
一部の研究者からはすでに教科書や論文で新名称を紹介したいという要望が出ており、peri-germ cell membraneという用語は速やかに世界に波及するでしょう。これまで問題だった呼称の混乱も解消し、今回の成果が植物科学の教育や研究に与える影響は小さくありません。精細胞と栄養細胞の境界に位置するインターフェースとして、peri-germ cell membraneは核酸やタンパク質の輸送や、花粉管における精細胞の先端輸送、精細胞の活性化などさまざまな受精過程に働くと推測されています。現時点でそれらは未解明ですが、名称統一を契機に新規参入する研究者が増えることで今後の研究の加速が期待されます。
イチョウやソテツの精子発見に象徴されるように、日本は植物生殖分野において重要な役割を果たしてきました[11,12]。本成果でも、国内外の多様な研究機関に属する日本人研究者が数多く名を連ねています。その背景には、母国語で教育や研究ができるように専門用語を整備し、先端の科学コミュニティを維持してきた我が国の学術的蓄積があります。精細胞を覆う膜にも内部(原)形質膜という日本語の呼称がありました。今後、原形質膜の意を含まずにperi-germ cell membraneと対応する日本語の新名称が専門家の協議で定められる予定です。新名称の発表を機に、先人達の築いてきた植物科学の価値や魅力が伝わることを願います。
研究費
本研究は、JSPS科研費(JP20H05778, JP20H05781, JP23K14214, JP24KJ0184)、ながひさ科学振興財団研究奨励金(G2023-01)、French National Research Agency(ANR-19-CE20-0012)、Sélection Végétale Avancée research program、National Research Agency France 2030 program、Association Nationale de la Recherche Technique (ANRT) PhD特別研究員制度(no. 2023/1284)の支援を受けて実施されました。
論文情報
タイトル: The peri-germ cell membrane: poorly characterized but key interface
for plant reproduction(日本語訳:Peri-germ cell membrane: 謎多き植物生殖のキーインターフェース)
著者: Naoya Sugi, Andrea R. M. Calhau, Nathanaël M. A. Jacquier, Marina Millan-Blanquez, Jörg D. Becker, Kevin Begcy, Frédéric Berger, Cécile Bousquet-Antonelli, Daniel Bouyer, Giampiero Cai, Alice Y. Cheung, Sílvia Coimbra, Philipp Denninger, Thomas Dresselhaus, José A. Feijó, John E. Fowler, Danny Geelen, Ueli Grossniklaus, Tetsuya Higashiyama, David Honys, Tomoko Igawa, Gwyneth Ingram, Yvon Jaillais, Mark A. Johnson, Mariko Kato, Miki Kawachi, Tomokazu Kawashima, Yu-Jin Kim, Hong-Ju Li, Sébastien Mongrand, Kazuki Motomura, Shiori Nagahara, Kohdai P. Nakajima, Brad Nelms, Li-Jia Qu, Arp Schnittger, Stefan Scholten, Stefanie Sprunck, Meng-Xiang Sun, David Twell, Dolf Weijers, Wei-Cai Yang, Daisuke Maruyama, Thomas Widiez
掲載雑誌: Nature Plants
DOI:https://doi.org/10.1038/s41477-024-01818-5
[画像4]https://digitalpr.jp/simg/1706/97072/150_117_2024101710442167106bf5034bc.png
[画像5]https://digitalpr.jp/simg/1706/97072/450_80_2024101710443067106bfec0600.jpg
用語説明
*1 三細胞性の花粉:栄養細胞の内部に2個の精細胞が存在する成熟花粉を三細胞性花粉とよびます。その他にも種によっては、精細胞形成前の雄原細胞の状態で花粉が成熟するニ細胞性花粉が作られます。このタイプの花粉は受粉後、花粉管の伸長中に雄原細胞が分裂して2個の精細胞を形成します。様々な植物の調査の結果、おおよそ7割の被子植物がニ細胞性の花粉を作るといわれています[13]。
参考文献など
1.Giménez-Martín, G., Risueño, M. C. & López-Sáez, J. F., (1969) Generative cell envelope in pollen grains as a secretion system, a postulate. Protoplasma 67: 223–235. https://doi.org/10.1007/BF01248742
2.Russell, S. D. & Cass, D. D., (1981) Ultrastructure of the sperms of Plumbago zeylanica. Protoplasma 107: 85–107. https://doi.org/10.1007/BF01275610
3.Dumas, C., Knox, R. B., McConchie, C. A. & Russell, S. D., (1984) Emerging physiological concepts in fertilization. What’s New Plant Physiol. 15: 17–20.
4.Theunis, C. H., McConchie, C. A. & Knox, R. B., (1985) Three-dimensional reconstruction of the generative cell and its wall connection in mature bicellular pollen of Rhododendron. Micron Microsc. Acta 16: 225–231. https://doi.org/10.1016/0739-6260(85)90047-4
5.Dumas, C., Knox, R. B. & Gaude, T., (1985) The spatial association of the sperm cells and vegetative nucleus in the pollen grain of Brassica. Protoplasma 124: 168–174. https://doi.org/10.1007/BF01290767
6.McConchie, C. A., Hough, T. & Knox, R. B., (1987) Ultrastructural analysis of the sperm cells of mature pollen of maize, Zea mays. Protoplasma 139: 9–19. https://doi.org/10.1007/BF01417530
7.Li, S., Zhou, L.-Z., Feng, Q.-N., McCormick, S. & Zhang, Y., (2013) The C-Terminal Hypervariable Domain Targets Arabidopsis ROP9 to the Invaginated Pollen Tube Plasma Membrane. Mol. Plant 6: 1362–1364. https://doi.org/10.1093/mp/sst098
8.Gilles, L. M. et al., (2021) Lipid anchoring and electrostatic interactions target NOT-LIKE-DAD to pollen endo-plasma membrane. J. Cell Biol. 220: e202010077. https://doi.org/10.1083/jcb.202010077
9.Billey,E. et al., (2021) LARP6C orchestrates posttranscriptional reprogramming of gene expression during hydration to promote pollen tube guidance. Plant Cell 33: 2637–2661. https://doi.org/10.1093/plcell/koab131
10.Wudick, M. M. et al., (2018) CORNICHON sorting and regulation of GLR channels underlie pollen tube Ca2+ homeostasis. Science 360: 533–536. https://doi.org/10.1126/science.aar6464
11.平瀬作五郎(1896)いてふノ精虫ニ就テ 植物学雑誌 第116号: 325–328. https://doi.org/10.15281/jplantres1887.10.116_325
12.池野成一郎 (1896) そてつノ精虫 植物学雑誌 第117号: 367-368. https://doi.org/10.15281/jplantres1887.10.117_367
13.Brewbaker, J. B. (1957) Pollen cytology and self-incompatibility systems in plants. J. Heredity 48: 271–277. https://doi.org/10.1093/oxfordjournals.jhered.a106742
プレスリリース詳細へ https://digitalpr.jp/r/97072
学問において、専門用語は円滑な教育研究活動の基盤となる必須の道具です。以降、世界中の教科書や論文で本名称が使われることからも、今回の名称統一は植物科学の重要な過程といえます。今後、日本語の新名称が専門家の協議で定められる予定です。
本成果は、植物科学誌「Nature Plants」に掲載されました(2024年10月15日)。
研究成果のポイント
精細胞を覆う生体膜に複数の呼称があることで植物生殖研究に混乱が生じていた。
世界中の研究者との協議で精細胞を覆う膜をperi-germ cell membraneと命名。
名称を統一したことで今後の重複受精の研究と教育に重要な貢献をした。
[画像1]https://digitalpr.jp/simg/1706/97072/450_248_20241016141217670f4b3170c37.jpg
図1 三細胞性花粉の模式図
これまで、文献によって精細胞の外側を覆う膜の呼びかたが異なっていたが、今回、世界の専門家の同意を得て「Peri-germ cell membrane」という名称に統一した
研究背景
植物の種子形成は、2つの精細胞が卵細胞と中央細胞という雌の細胞を同時に受精させる重複受精によって始まります。2つの精細胞は花粉や花粉管の栄養細胞の内部で、一重膜に覆われた状態で存在します(図1)。この膜は重複受精で重要な役割を担っているにも関わらず、統一された名称をもたず、文献によってさまざまな形で呼ばれてきました。
シロイヌナズナやイネ、トウモロコシなどの植物(被子植物)で見られる三細胞性の花粉*1は、雄しべの葯(やく)で小胞子が2回の分裂をすることで作られます。まず、最初の細胞分裂で小さめの雄原細胞と栄養細胞が作られると、栄養細胞は雄原細胞を中に取り込みます。続く細胞分裂で雄原細胞から2個の精細胞が作られて花粉は成熟します(図2)。細胞の取り込みという特徴的な過程を経るため、雄原細胞や精細胞は栄養細胞の細胞膜に由来する一重膜に包み込まれた状態で存在します。この膜は古くから知られており、1969年にはすでに雄原細胞を覆う膜を意味するgenerative cell envelopeと記述されています[1]。ところがその後、internal plasma membrane of the vegetative cell、inner plasma membrane of the pollen grain、inner vegetative cell plasma membrane、pollen endo-plasma membraneなど、この膜はさまざまな名称で呼ばれることとなってきました[2-6]。近年、精細胞を覆う膜が重複受精に果たす役割が議論されるようになり、乱立した専門用語を統一する機運が徐々に高まってきました。
[画像2]https://digitalpr.jp/simg/1706/97072/450_232_20241016141217670f4b31865b6.jpg
図2 三細胞性花粉の花粉形成過程
研究内容
名称の統一に向けては、精細胞を覆う膜の性質を適切に表し、かつ、簡潔で使いやすいものが求められました。しかし、花粉や重複受精をさまざまな観点から扱う世界中の専門家の意見を取り入れ、皆が納得する名称を提案する作業は大きな挑戦でした。そこでThomas Widiez博士と丸山大輔准教授が着目したのは、旧名称のほとんどが細胞膜(原形質膜)を意味するplasma membraneを含む点でした。精細胞を覆う膜は確かに栄養細胞の細胞膜を起源としていますが、近年の研究でこの膜に含まれるリン脂質やタンパク質の構成が通常の細胞膜とは異なることがわかってきています[7-10]。そこで、典型的な3種の細胞膜タンパク質SYP121、 SYP132、 LTI6bを蛍光タンパク質でラベルして、その局在を確認しました。すると、いずれも花粉管栄養細胞の細胞膜には存在する一方、精細胞を覆う膜では検出されないことが示されました(図3)。これは、今回議論となっている精細胞を覆う膜が典型的な細胞膜とは異なる性質を持つことを意味しています。これにより、新名称から“plasma membrane”を除くことで合意が得られました。その後の議論で、雄原細胞と精細胞の両方の意が含まれるgerm cellという語に対して、「周辺の、覆う」の意をもつ接頭辞のperiを合わせたperi-germ cell membraneが新名称として選ばれました。
[画像3]https://digitalpr.jp/simg/1706/97072/450_269_20241016141230670f4b3e9dada.jpg
図3 典型的な細胞膜タンパク質はperi-germ cell membraneに局在しない
蛍光タンパク質でラベルした典型的な細胞膜タンパク質(SYP121, SYP132, LTI6b)とperi-germ cell membrane局在シグナル(Lyn24)をつないだ蛍光タンパク質を花粉管栄養細胞で発現させた。伸長中の花粉管において典型的な細胞膜タンパク質は花粉管栄養細胞膜への局在を示した。一方で、伸長中の花粉管および精細胞放出後の両方においてperi-germ cell membraneには局在しなかった。赤い矢印で示した部分の相対蛍光強度をグラフに示している
今後の展開
一部の研究者からはすでに教科書や論文で新名称を紹介したいという要望が出ており、peri-germ cell membraneという用語は速やかに世界に波及するでしょう。これまで問題だった呼称の混乱も解消し、今回の成果が植物科学の教育や研究に与える影響は小さくありません。精細胞と栄養細胞の境界に位置するインターフェースとして、peri-germ cell membraneは核酸やタンパク質の輸送や、花粉管における精細胞の先端輸送、精細胞の活性化などさまざまな受精過程に働くと推測されています。現時点でそれらは未解明ですが、名称統一を契機に新規参入する研究者が増えることで今後の研究の加速が期待されます。
イチョウやソテツの精子発見に象徴されるように、日本は植物生殖分野において重要な役割を果たしてきました[11,12]。本成果でも、国内外の多様な研究機関に属する日本人研究者が数多く名を連ねています。その背景には、母国語で教育や研究ができるように専門用語を整備し、先端の科学コミュニティを維持してきた我が国の学術的蓄積があります。精細胞を覆う膜にも内部(原)形質膜という日本語の呼称がありました。今後、原形質膜の意を含まずにperi-germ cell membraneと対応する日本語の新名称が専門家の協議で定められる予定です。新名称の発表を機に、先人達の築いてきた植物科学の価値や魅力が伝わることを願います。
研究費
本研究は、JSPS科研費(JP20H05778, JP20H05781, JP23K14214, JP24KJ0184)、ながひさ科学振興財団研究奨励金(G2023-01)、French National Research Agency(ANR-19-CE20-0012)、Sélection Végétale Avancée research program、National Research Agency France 2030 program、Association Nationale de la Recherche Technique (ANRT) PhD特別研究員制度(no. 2023/1284)の支援を受けて実施されました。
論文情報
タイトル: The peri-germ cell membrane: poorly characterized but key interface
for plant reproduction(日本語訳:Peri-germ cell membrane: 謎多き植物生殖のキーインターフェース)
著者: Naoya Sugi, Andrea R. M. Calhau, Nathanaël M. A. Jacquier, Marina Millan-Blanquez, Jörg D. Becker, Kevin Begcy, Frédéric Berger, Cécile Bousquet-Antonelli, Daniel Bouyer, Giampiero Cai, Alice Y. Cheung, Sílvia Coimbra, Philipp Denninger, Thomas Dresselhaus, José A. Feijó, John E. Fowler, Danny Geelen, Ueli Grossniklaus, Tetsuya Higashiyama, David Honys, Tomoko Igawa, Gwyneth Ingram, Yvon Jaillais, Mark A. Johnson, Mariko Kato, Miki Kawachi, Tomokazu Kawashima, Yu-Jin Kim, Hong-Ju Li, Sébastien Mongrand, Kazuki Motomura, Shiori Nagahara, Kohdai P. Nakajima, Brad Nelms, Li-Jia Qu, Arp Schnittger, Stefan Scholten, Stefanie Sprunck, Meng-Xiang Sun, David Twell, Dolf Weijers, Wei-Cai Yang, Daisuke Maruyama, Thomas Widiez
掲載雑誌: Nature Plants
DOI:https://doi.org/10.1038/s41477-024-01818-5
[画像4]https://digitalpr.jp/simg/1706/97072/150_117_2024101710442167106bf5034bc.png
[画像5]https://digitalpr.jp/simg/1706/97072/450_80_2024101710443067106bfec0600.jpg
用語説明
*1 三細胞性の花粉:栄養細胞の内部に2個の精細胞が存在する成熟花粉を三細胞性花粉とよびます。その他にも種によっては、精細胞形成前の雄原細胞の状態で花粉が成熟するニ細胞性花粉が作られます。このタイプの花粉は受粉後、花粉管の伸長中に雄原細胞が分裂して2個の精細胞を形成します。様々な植物の調査の結果、おおよそ7割の被子植物がニ細胞性の花粉を作るといわれています[13]。
参考文献など
1.Giménez-Martín, G., Risueño, M. C. & López-Sáez, J. F., (1969) Generative cell envelope in pollen grains as a secretion system, a postulate. Protoplasma 67: 223–235. https://doi.org/10.1007/BF01248742
2.Russell, S. D. & Cass, D. D., (1981) Ultrastructure of the sperms of Plumbago zeylanica. Protoplasma 107: 85–107. https://doi.org/10.1007/BF01275610
3.Dumas, C., Knox, R. B., McConchie, C. A. & Russell, S. D., (1984) Emerging physiological concepts in fertilization. What’s New Plant Physiol. 15: 17–20.
4.Theunis, C. H., McConchie, C. A. & Knox, R. B., (1985) Three-dimensional reconstruction of the generative cell and its wall connection in mature bicellular pollen of Rhododendron. Micron Microsc. Acta 16: 225–231. https://doi.org/10.1016/0739-6260(85)90047-4
5.Dumas, C., Knox, R. B. & Gaude, T., (1985) The spatial association of the sperm cells and vegetative nucleus in the pollen grain of Brassica. Protoplasma 124: 168–174. https://doi.org/10.1007/BF01290767
6.McConchie, C. A., Hough, T. & Knox, R. B., (1987) Ultrastructural analysis of the sperm cells of mature pollen of maize, Zea mays. Protoplasma 139: 9–19. https://doi.org/10.1007/BF01417530
7.Li, S., Zhou, L.-Z., Feng, Q.-N., McCormick, S. & Zhang, Y., (2013) The C-Terminal Hypervariable Domain Targets Arabidopsis ROP9 to the Invaginated Pollen Tube Plasma Membrane. Mol. Plant 6: 1362–1364. https://doi.org/10.1093/mp/sst098
8.Gilles, L. M. et al., (2021) Lipid anchoring and electrostatic interactions target NOT-LIKE-DAD to pollen endo-plasma membrane. J. Cell Biol. 220: e202010077. https://doi.org/10.1083/jcb.202010077
9.Billey,E. et al., (2021) LARP6C orchestrates posttranscriptional reprogramming of gene expression during hydration to promote pollen tube guidance. Plant Cell 33: 2637–2661. https://doi.org/10.1093/plcell/koab131
10.Wudick, M. M. et al., (2018) CORNICHON sorting and regulation of GLR channels underlie pollen tube Ca2+ homeostasis. Science 360: 533–536. https://doi.org/10.1126/science.aar6464
11.平瀬作五郎(1896)いてふノ精虫ニ就テ 植物学雑誌 第116号: 325–328. https://doi.org/10.15281/jplantres1887.10.116_325
12.池野成一郎 (1896) そてつノ精虫 植物学雑誌 第117号: 367-368. https://doi.org/10.15281/jplantres1887.10.117_367
13.Brewbaker, J. B. (1957) Pollen cytology and self-incompatibility systems in plants. J. Heredity 48: 271–277. https://doi.org/10.1093/oxfordjournals.jhered.a106742
プレスリリース詳細へ https://digitalpr.jp/r/97072
提供:




